Protein ubiquitination
收藏reactome.org2025-01-15 收录
下载链接:
https://reactome.org/PathwayBrowser/#/R-HSA-8852135
下载链接
链接失效反馈官方服务:
资源简介:
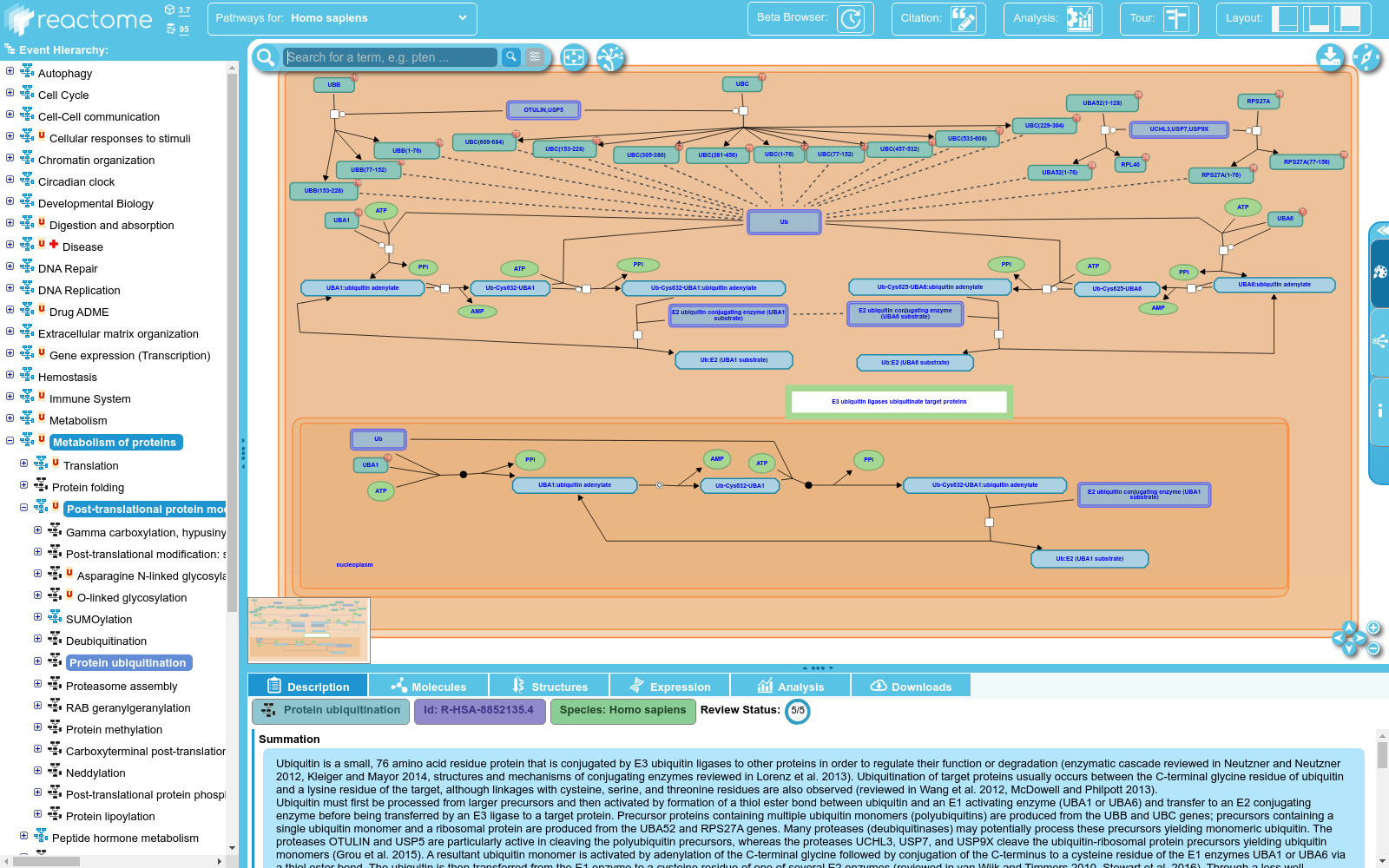
Ubiquitin is a small, 76 amino acid residue protein that is conjugated by E3 ubiquitin ligases to other proteins in order to regulate their function or degradation (enzymatic cascade reviewed in Neutzner and Neutzner 2012, Kleiger and Mayor 2014, structures and mechanisms of conjugating enzymes reviewed in Lorenz et al. 2013). Ubiquitination of target proteins usually occurs between the C-terminal glycine residue of ubiquitin and a lysine residue of the target, although linkages with cysteine, serine, and threonine residues are also observed (reviewed in Wang et al. 2012, McDowell and Philpott 2013).<br>Ubiquitin must first be processed from larger precursors and then activated by formation of a thiol ester bond between ubiquitin and an E1 activating enzyme (UBA1 or UBA6) and transfer to an E2 conjugating enzyme before being transferred by an E3 ligase to a target protein. Precursor proteins containing multiple ubiquitin monomers (polyubiquitins) are produced from the UBB and UBC genes; precursors containing a single ubiquitin monomer and a ribosomal protein are produced from the UBA52 and RPS27A genes. Many proteases (deubiquitinases) may potentially process these precursors yielding monomeric ubiquitin. The proteases OTULIN and USP5 are particularly active in cleaving the polyubiquitin precursors, whereas the proteases UCHL3, USP7, and USP9X cleave the ubiquitin-ribosomal protein precursors yielding ubiquitin monomers (Grou et al. 2015). A resultant ubiquitin monomer is activated by adenylation of the C-terminal glycine followed by conjugation of the C-terminus to a cysteine residue of the E1 enzymes UBA1 or UBA6 via a thiol ester bond. The ubiquitin is then transferred from the E1 enzyme to a cysteine residue of one of several E2 enzymes (reviewed in van Wijk and Timmers 2010, Stewart et al. 2016). Through a less well characterized mechanism, E3 ubiquitin ligases then bring a target protein and the E2-ubiquitin conjugate into proximity so that the ubiquitin is transferred via formation of an amide bond to a particular lysine residue (or, in rarer cases, a thiol ester bond to a cysteine residue or an ester bond to a serine or threonine residue) of the target protein (reviewed in Berndsen and Wolberger 2014). Based on protein homologies, families of E3 ubiquitin ligases have been identified that include RING-type ligases (reviewed in Deshaies et al. 2009, Metzger et al. 2012, Metzger et al. 2014), HECT-type ligases (reviewed in Rotin et al. 2009, Metzger et al. 2012), and RBR-type ligases (reviewed in Dove et al. 2016). A subset of the RING-type ligases participate in CULLIN-RING ligase complexes (CRLs which include SCF complexes, reviewed in Lee and Zhou 2007, Genschik et al. 2013, Skaar et al. 2013, Lee et al. 2014).<br>Some E3-E2 combinations catalyze mono-ubiquitination of the target protein (reviewed in Nakagawa and Nakayama 2015). Other E3-E2 combinations catalyze conjugation of further ubiquitin monomers to the initial ubiquitin, forming polyubiquitin chains. (It may also be possible for some E3-E2 combinations to preassemble polyubiquitin and transfer it as a unit to the target protein.) Ubiquitin contains several lysine (K) residues and a free alpha amino group to which further ubiquitin can be conjugated. Thus different types of polyubiquitin are possible: K11 linked polyubiquitin is observed in endoplasmic reticulum-associated degradation (ERAD), K29 linked polyubiquitin is observed in lysosomal degradation, K48 linked polyubiquitin directs target proteins to the proteasome for degradation, whereas K63 linked polyubiquitin generally acts as a scaffold to recruit other proteins in several cellular processes, notably DNA repair (reviewed in Komander et al. 2009). Ubiquitination is highly regulated (reviewed in Vittal et al. 2015) and affects all cellular processes including DNA damage response (reviewed in Brown and Jackson 2015), immune signaling (reviewed in Park et al. 2014, Lutz-Nicoladoni et al. 2015), and regulation of normal and cancerous cell growth (reviewed in Skaar and Pagano 2009, Yerlikaya and Yontem 2013, Strikoudis et al. 2014).
泛素是一种由76个氨基酸残基组成的微小蛋白质,通过E3泛素连接酶与其他蛋白质共价结合,以调节其功能或降解(酶促级联反应详见Neutzner和Neutzner 2012,Kleiger和Mayor 2014,连接酶的结构与机制详见Lorenz等2013年的综述)。泛素化通常发生在泛素的C端甘氨酸残基与靶蛋白的赖氨酸残基之间,尽管也有与半胱氨酸、丝氨酸和苏氨酸残基的连接(详见Wang等2012年,McDowell和Philpott 2013年的综述)。泛素首先需从较大的前体蛋白中加工处理,并通过泛素与E1激活酶(UBA1或UBA6)之间形成硫酯键来激活,然后转移至E2连接酶,最后由E3连接酶转移至靶蛋白。含有多个泛素单体(多泛素)的前体蛋白由UBB和UBC基因产生;含有单个泛素单体和核糖体蛋白的前体蛋白由UBA52和RPS27A基因产生。许多蛋白酶(去泛素化酶)可能将这些前体蛋白加工处理,产生单体泛素。OTULIN和USP5蛋白酶在切割多泛素前体蛋白方面尤为活跃,而UCHL3、USP7和USP9X蛋白酶则切割泛素-核糖体蛋白前体蛋白,产生泛素单体(详见Grou等2015年的研究)。产生的单体泛素通过C端甘氨酸的腺苷化激活,随后通过硫酯键将其C端连接到E1酶UBA1或UBA6的半胱氨酸残基。随后,泛素从E1酶转移至多个E2酶之一的半胱氨酸残基(详见van Wijk和Timmers 2010年,Stewart等2016年的综述)。通过一种尚未充分表征的机制,E3泛素连接酶将靶蛋白和E2-泛素连接物拉近,以便通过形成酰胺键将泛素转移至靶蛋白的特定赖氨酸残基(或,在罕见情况下,通过硫酯键连接到半胱氨酸残基,或通过酯键连接到丝氨酸或苏氨酸残基)。基于蛋白质同源性,已鉴定出包括RING型连接酶(详见Deshaies等2009年,Metzger等2012年,Metzger等2014年的综述)、HECT型连接酶(详见Rotin等2009年,Metzger等2012年的综述)和RBR型连接酶(详见Dove等2016年的综述)在内的E3泛素连接酶家族。其中,一些RING型连接酶参与CULLIN-RING连接酶复合物(CRLs,包括SCF复合物,详见Lee和Zhou 2007年,Genschik等2013年,Skaar等2013年,Lee等2014年的综述)。某些E3-E2组合催化靶蛋白的单泛素化(详见Nakagawa和Nakayama 2015年的综述)。其他E3-E2组合催化进一步泛素单体与初始泛素共价连接,形成多泛素链。(某些E3-E2组合预先组装多泛素并将其作为一个单元转移到靶蛋白上也是可能的。)泛素含有多个赖氨酸(K)残基和一个自由的α-氨基,可以进一步连接其他泛素。因此,可能存在不同类型的泛素化:K11连接的多泛素在质膜内质网相关降解(ERAD)中观察到,K29连接的多泛素在溶酶体降解中观察到,K48连接的多泛素将靶蛋白导向蛋白酶体进行降解,而K63连接的多泛素通常作为支架,在多种细胞过程中招募其他蛋白质,尤其是在DNA修复过程中(详见Komander等2009年的综述)。泛素化受到高度调控(详见Vittal等2015年的综述),并影响包括DNA损伤反应(详见Brown和Jackson 2015年的综述)、免疫信号(详见Park等2014年,Lutz-Nicoladoni等2015年的综述)以及正常和癌变细胞生长的调控(详见Skaar和Pagano 2009年,Yerlikaya和Yontem 2013年,Strikoudis等2014年的综述)。
提供机构:
Reactome
搜集汇总
数据集介绍
背景与挑战
背景概述
该数据集详细描述了人类(Homo sapiens)蛋白质泛素化的生物过程,涵盖了从泛素前体加工、E1/E2/E3酶级联反应到目标蛋白修饰的完整机制。其特点包括解释了单泛素化和多聚泛素化(如K48、K63链接)的不同功能,并关联了该过程在DNA损伤响应、免疫调控和细胞生长等关键细胞通路中的作用。数据集经过全面审核(5/5分),提供了系统的分子机制概述和相关参考文献。
以上内容由遇见数据集搜集并总结生成


